
There exists a threshold beyond which bone loses its remarkable self-repair capacity. The critical-size defect — that gap which does not spontaneously reconstitute over the organism's lifespan and which, left to itself, results in pseudoarthrosis or fibrous colonization — represents the true battleground of bone regeneration. The causes are well known and clinically significant: oncological resections, high-energy trauma with substance loss, infections requiring extensive debridement, osteosynthesis failures, prosthetic revisions with periprosthetic osteolysis. In all these scenarios, physiological healing — which proceeds through embryological recapitulation via the inflammatory cascade, callus formation, and subsequent remodeling — comes to a halt because the void to be filled exceeds the capacity of endogenous mechanisms.
The gold standard remains autograft, ideally osteoconductive, osteoinductive, and osteogenic all at once, but the biological cost is considerable: donor site morbidity, chronic pain at the harvest region, intrinsically limited tissue quantity, and a geometry that rarely matches the defect to be reconstructed. Allograft alleviates the availability problem but introduces immunogenicity, risk of pathogen transmission, and, most critically, an almost complete loss of osteoinductive potential due to sterilization and devitalization processes. It is within this fracture between clinical need and the limitations of native biological material that tissue engineering has inserted its project: to build synthetic or natural scaffolds capable of guiding new bone formation, using materials that engage in dialogue with tissue biology rather than merely filling space. Silk fibroin is, in this dialogue, one of the most convincing interlocutors.
Why fibroin: molecular architecture and biomimetic rationale
The strength of fibroin as a biomaterial resides in an apparent contradiction: it is a structural polymer produced by an invertebrate, yet it behaves with vertebrate tissues in a way that few synthetic materials achieve. Bombyx mori fibroin is organized into a heavy chain of approximately 390 kDa and a light chain of approximately 26 kDa, linked by a disulfide bridge, with the glycoprotein P25 non-covalently associated. However, it is the heavy chain sequence that determines its properties: highly repetitive hydrophobic blocks, dominated by (Gly-Ala-Gly-Ala-Gly-Ser) motifs, which collapse into crystalline β-sheets, alternating with more hydrophilic and less ordered amorphous regions. This alternation between crystalline and amorphous domains generates a semicrystalline material in which β-sheets act as physical crosslinking nodes, conferring tensile strength and stiffness, while the amorphous regions provide elasticity and energy absorption capacity.
Conformational control is what makes fibroin so versatile for scaffold design. The protein can assume the silk I state, metastable and water-soluble, and the silk II state, rich in β-sheets, thermodynamically stable and insoluble. The transition between the two, inducible via alcohol treatment, water annealing, sonication, shear, or pH changes, is the true control panel of the material: increasing the β-sheet fraction stiffens the structure and slows its degradation, while reducing it yields softer, faster-resorbing matrices. Added to this is a plurality of processing routes, from aqueous solution to organic solvents such as hexafluoroisopropanol, enabling the production of films, sponges, hydrogels, electrospun fibers, and three-dimensional printing inks from the same raw material.
On the biological level, the removal of sericin during degumming is the necessary condition for fibroin to express its biocompatibility: sericin, in fact, is the sensitizing component of silk, and its residual persistence historically explains the adverse reactions attributed to silk sutures in the past. Purified fibroin, conversely, elicits a contained inflammatory response, supports the adhesion and proliferation of numerous cell lines, and degrades into metabolizable peptides and amino acids. The biomimetic rationale arises precisely here: fibroin can play, with respect to the mineral component of bone, a role analogous to that of type I collagen in the native matrix, offering a fibrous organic template capable of organizing calcium phosphate deposition.
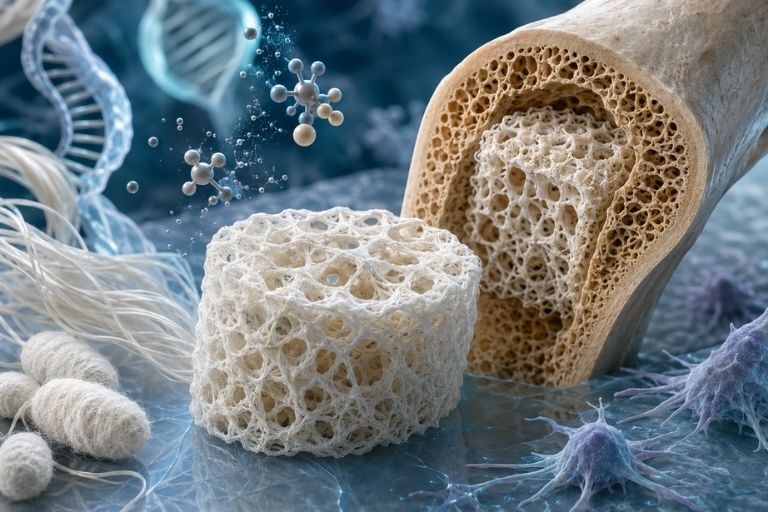
Reconstructing the bone matrix: silk-guided mineralization
Bone is a hierarchical nanocomposite in which an organic matrix of type I collagen is permeated by carbonated hydroxyapatite crystals, calcium-deficient and nanometric in size, aligned along the axis of the collagen fibrils. Reproducing this intimate interpenetration between organic and mineral phases is the condition for a scaffold not merely to fill a void but to behave as a true analogue of the bone extracellular matrix. Fibroin lends itself to this task because its amino acid residues — particularly the carboxyl groups of acidic amino acids and the hydroxyl groups of serine — can serve as heterogeneous nucleation sites for calcium phosphate precipitation.
Mineralization strategies follow several complementary pathways. Immersion in simulated body fluid induces superficial apatitic deposition that mimics the composition of bone mineral, exploiting solution supersaturation and the nucleating action of the protein surface. Coprecipitation, in which crystal formation occurs in situ during gelation or solidification of the material, allows the mineral phase to be distributed throughout the entire scaffold volume rather than confined to the surface. The polymer-induced liquid precursor approach introduces a fluid amorphous phase that infiltrates the finest pores of the matrix and mineralizes from within, more faithfully reproducing the manner in which mineralization occurs physiologically within and between collagen fibrils.
A decisive step concerns the chemical functionalization of fibroin to amplify its nucleating capacity. The introduction of phosphate groups, for example through phosphorylation reactions that mimic the presence of non-collagenous bone phosphoproteins, significantly increases the density of nucleation sites and accelerates the formation of well-organized apatite. The outcome of these strategies is not merely a stiffer material but a surface chemically recognized by osteoprogenitor cells as an osteoconductive environment, capable of triggering the adhesion, orientation, and subsequent differentiation of recruited cells.
Scaffold architecture and fabrication technologies
No surface chemistry, however sophisticated, compensates for inadequate architecture. Bone regeneration requires interconnected porosity with pore diameters typically ranging from one hundred to four hundred microns: pores too small hinder cell infiltration and, above all, vascular penetration; pores too large sacrifice available surface area for adhesion and compromise mechanical strength. Interconnection between pores is at least as crucial as their size, because it determines whether vessels and cells colonize the entire volume rather than halting at peripheral regions.
Fabrication technologies allow these parameters to be modulated with considerable precision. Porogen leaching, in which salt crystals of controlled size are dispersed in the fibroin solution and then leached out, yields predictable and easily tunable porosity by acting on crystal size and volume fraction. Freeze-drying exploits ice crystal sublimation to generate porous networks, and when freezing is directional, it produces anisotropic architectures with aligned channels reminiscent of the osteonal organization of cortical bone. Electrospinning generates nanofibrous mats that dimensionally mimic extracellular matrix fibrils, offering a topographically affine substrate to that which cells encounter in native tissue. Three-dimensional printing and bioprinting, finally, have shifted the discipline's center of gravity toward patient-specific fabrication: starting from tomographic imaging, it is possible to design scaffolds whose external geometry matches the defect and whose internal microarchitecture is designed to simultaneously optimize porosity, vascularization, and mechanical performance.
Fibroin adapts to all these routes with a rare plasticity, and its ability to consolidate through the transition to the silk II state offers a stabilization mechanism that requires no toxic chemical crosslinkers. Sponges, injectable hydrogels, films, nanofibers, and printed constructs thus derive from the same protein solution, selected case by case according to the anatomical site and expected mechanical load.
The mechanical question
Bone is not mechanically homogeneous, and this heterogeneity imposes on the scaffold a tuning requirement that is difficult to satisfy. Trabecular bone exhibits elastic moduli on the order of hundreds of megapascals, while cortical bone reaches gigapascals: a matrix designed for cancellous bone is mechanically inadequate to support loads in a diaphyseal segment, and an overly stiff matrix introduces the opposite risk of stress shielding, where mechanical protection deprives the newly formed bone of the stimuli necessary for remodeling and promotes its resorption.
Pure fibroin, though strong among natural polymers, rarely achieves by itself the values required for load-bearing sites. The most established solution is the composite approach, in which the protein matrix is reinforced with osteoconductive inorganic phases: hydroxyapatite, beta-tricalcium phosphate, bioactive glasses. These reinforcements not only elevate the compressive modulus and strength of the scaffold, bringing them closer to values of the host bone, but also add their own bioactive contribution, because their gradual resorption releases calcium and phosphate ions that fuel the mineralization of the forming tissue. The dosage of the inorganic phase thus becomes a design parameter with dual valence, mechanical and biological, and its balancing with the β-sheet fraction of the protein component allows a property gradient to be built that, ideally, accompanies the transition from the initial stiffness of the scaffold to the progressively acquired stiffness of the bone that replaces it.
Bioactive functionalization and growth factor delivery
An osteoconductive scaffold guides bone growth along its surfaces, but reconstruction of larger defects requires active osteoinduction, i.e., the recruitment and differentiation of osteoprogenitor cells. Fibroin offers for this purpose a valuable quality that is less obvious than it seems: it is an excellent vehicle for the controlled release of bioactive molecules. Its semicrystalline structure protects encapsulated growth factors from denaturation, and release kinetics can be modulated by regulating the β-sheet content, which acts as a diffusional barrier and a regulator of matrix degradation.
Bone morphogenetic protein 2 is the paradigm of this application: incorporated into a fibroin matrix, it is released in a sustained manner and retains its bioactivity longer than with less protective carriers, reducing the recourse to supraphysiological doses that, in clinical practice, have raised concerns regarding side effects such as ectopic bone formation and edema. Alongside growth factors, functionalization with adhesion sequences, foremost the Arg-Gly-Asp tripeptide recognized by integrins, enhances cell anchoring and transduction of mechanical signals that direct osteogenic fate. Fibroin can also deliver ions and active agents with additional therapeutic value, from strontium and magnesium ions, which promote osteoblastic activity and modulate osteoclastic activity, to antibacterial agents designed for defects at risk of infection, transforming the scaffold from a simple structural support into an integrated local delivery system.
The challenge of vascularization
The most stubborn frontier of bone regeneration is not matrix deposition but blood supply. Beyond a distance on the order of a few hundred microns from the nearest vessel, oxygen and nutrient diffusion becomes insufficient, and the center of a large scaffold undergoes hypoxia and necrosis before new bone formation can consolidate. Regeneration of a critical defect is therefore, even before an osteogenic problem, an angiogenic problem: without a vascular network that rapidly penetrates the entire construct, every osteoinductive strategy remains confined to the periphery.
Responses to this challenge are multiple and often combined. On the biological side, recourse is made to the release of vascular endothelial growth factor, sometimes in temporal sequence coordinated with osteogenic signals to reproduce the natural precedence of angiogenesis over osteogenesis, and to co-culture of osteoprogenitor and endothelial cells to trigger in vitro the formation of primordial vascular structures. On the architectural side, channels and preformed porosity networks are designed to act as guides for vascular invasion, exploiting precisely the geometric freedom offered by three-dimensional printing of fibroin. The ability of the silk matrix to simultaneously support the release of pro-angiogenic signals and a microarchitecture that favors their effect renders fibroin a particularly suitable substrate for these prevascularization strategies, which likely constitute the real watershed between experimental success on small defects and clinical translation on the defects that matter.
Cellular interaction and osteogenic differentiation
The ultimate efficacy of a scaffold is measured by the behavior of the cells that colonize it. Mesenchymal stem cells, recruited from the marrow, periosteum, or deliberately introduced into the construct, respond to fibroin with adhesion, proliferation, and, under appropriate conditions, commitment toward the osteoblastic lineage. This commitment is reflected in the sequential expression of canonical markers: early activation of Runx2, the master transcriptional regulator of the osteogenic program; increase in alkaline phosphatase as a marker of the initial maturation phase; and finally deposition of osteocalcin, osteopontin, and type I collagen signaling the transition toward the mature phenotype and competence for mineralization.
What makes fibroin particularly interesting is the possibility to drive this differentiation through the physical properties of the material, in addition to biochemical signals. Substrate stiffness is a recognized determinant of mesenchymal cell fate, and the tunability of fibroin's elastic modulus, achievable by varying the β-sheet content or the fraction of inorganic reinforcement, offers a mechanotransductive lever to direct cells toward osteogenesis. Topography also matters: nanofibrous substrates obtained by electrospinning, reproducing the dimensional scale of the native matrix, influence cell morphology, cytoskeletal organization, and consequent activation of mechanosensitive signaling pathways. Fibroin, in other words, is not a passive support but an instructive material, which through its chemistry, stiffness, and geometry actively participates in the differentiative decision.
Proteolytic degradation and remodeling
An ideal scaffold is destined to disappear. Its function is transient: it must support and guide new bone formation and then progressively yield its place to regenerated tissue, avoiding persistence as a foreign body and interference with remodeling. The advantage of fibroin, in this respect, is that its degradation is proteolytic and therefore intrinsically biological: enzymes such as proteases and matrix metalloproteinases, recruited in the context of the tissue response, break down the protein into peptide fragments and amino acids that are reabsorbed and metabolized without generating the acidic products that characterize the degradation of certain synthetic polyesters and that can trigger local inflammation.
The challenge is not degradability per se, but its synchronization with new bone formation. A matrix that dissolves too rapidly collapses before the bone has acquired structural autonomy; a matrix that persists too long impedes remodeling and occupies the space that newly formed tissue should colonize. Control of β-sheet content is precisely the tool that allows this kinetics to be calibrated: increasing crystallinity slows enzymatic action and extends scaffold persistence, reducing it accelerates resorption. The possibility of constructing materials with degradation times designed to accompany the expected healing course in a specific anatomical site is one of the advantages that distinguishes fibroin from most other natural biomaterials, whose resorption is far less governable.
Preclinical evidence and clinical proximity
The accumulation of preclinical data outlines a coherent picture. In calvarial defect models — the classic paradigm for testing osteoconduction and osteoinduction — composite fibroin/hydroxyapatite scaffolds, especially when enriched with osteoinductive factors or osteoprogenitor cells, produce superior new bone formation compared to controls and progressive replacement of the material with vital bone. In segmental long bone defect models — a more challenging scenario due to mechanical loads and defect extent — composite and prevascularized strategies demonstrate the ability to support bone bridges across gaps that controls fail to close.
